Рибосома (от «РНК» и soma – тело) – клеточный немембранный органоид, осуществляющий трансляцию (считывание кода мРНК и синтез полипептидов). Молекула белка рождается в цитоплазме клетки на свободных рибосомах или на цистернах транспортно-накопительной системы. Специальные белки шапероны, укладывают растущую цепочку в ажурную конструкцию. Затем, если нужно, белок достраивают. Различают 2 основных типа рибосом – прокариотные и эукариотные. В митохондриях и хлоропластах также имеются рибосомы, которые близки к рибосомам прокариот. Рибосомы эукариот расположены на мембранах эндоплазматической сети (гранулярная ЭС) и в цитоплазме. Прикрепленные к мембранам рибосомы синтезируют белок «на экспорт», а свободные рибосомы – для нужд самой клетки.
Рибосомы прокариот
У прокариот генетический материал не изолирован от аппарата трансляции, и прокариотные рибосомы занимают почти весь цитоплазматический компартмент. Относительное (по сравнению с другими органеллами) количество рибосом у прокариотов выше, чем у эукариотов, и это обеспечивает более высокую активность их метаболизма, а также более высокую скорость их роста и размножения. Рибосомы - это множественные генеральные микрокомпартменты, которые находятся в цитоплазматическом компартменте и выполняют роль универсальной белоксинтезирующей органеллы. За исключением редких случаев, когда полипептиды синтезируются нерибосомным путем, аминокислоты связываются в линейную цепь только благодаря ферментативной активности рибосом.
Биосинтетический процесс трансляции уникален, поскольку информация о порядке расположения нуклеотидных триплетов (код мРНК) переводится в информацию о порядке расположения аминокислот (код полипептида). Посредником между этими кодами, или «адаптером» (англ. adapter - звукосниматель аудиосистемы) является тРНК. Она доставляет аминокислоту в пептидилтрансферазный центр и одновременно с этим распознает ее кодон в молекуле мРНК.
Типы рибосом. Рибосома - это мультимолекулярный комплекс, состоящий из рРНК и рибосомных белков в массовом соотношении 2:1. В рабочем состоянии рибосома, или «моносома» представляет собой частицу диаметром 25 нм, которая состоит из двух субъединиц - большой L-субъединицы (от англ. large) и малой S-субъединицы (от англ. small). Они имеют разный состав, разную морфологию и выполняют разные функции.
По количественным признакам все рибосомы подразделяются на два типа - прокариотный и эукариотный. Прокариотная рибосома имеет коэффициент седиментации 70S (субъединицы 50S и 30S), а эукариотная рибосома -80S (субъединицы 60S и 40S). Прокариотная рибосома содержит три молекулы рРНК - 23S (~3000 нуклеотидов), 16S (~1500 нуклеотидов) и 5S (~120 нуклеотидов), а также 53-65 однокопий-ных белков. Эукариотная рибосома устроена сложнее прокариотной. Она содержит не три, а четыре молекулы рРНК -28S (4000-6000 нуклеотидов), 18S (1750-1850 нуклеотидов), 5S (~120 нуклеотидов) и 5,8S (~150 нуклеотидов), а также более богатый набор однокопийных белков (70-84).
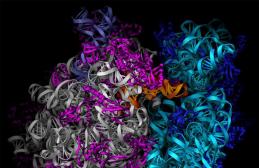
Механизм трансляции. Хотя современные представления об архитектуре рибосом и процессе трансляции сложились на основе данных, полученных на бактериях, доказано, что работа рибосомы универсальна у всех трех глобальных доменов. Результаты рентген-структурного анализа с уровнем разрешения 5,5А и криоэлектронной микроскопии дали для рибосомы Е. coli картину геометрического тела сложнейшей конфигурации, состоящего из взаимно переплетенных молекул рРНК и белка. Внутри него, а также на его поверхности имеются каналы, борозды, углубления, площадки, выступы и мостики.
Субстратами для биосинтеза полипептидной цепи служат аминоацил-тРНК, причем для каждой аминокислоты существует своя тРНК и своя аминоацил-тРНК-синтетаза. Специфические тРНК (~75 нуклеотидов) различаются по первичной структуре, однако все они имеют стандартную Г-образную третичную структуру. На дистальном конце длинного «локтя» находится антикодон, комплементарный триплету мРНК, который кодирует специфическую аминокислоту. На дистальном конце короткого «локтя» всех тРНК находится 3"-концевая последовательность ССА. К аденозину (по его 2"- или З"-гидроксильному радикалу) присоединяется а-карбоксильная группа специфической аминокислоты. В ходе трансляции антикодон длинного «локтя» распознает на S-субъединице кодон мРНК, а короткий «локоть» с аминокислотой взаимодействует на L-субъединице с пептидилтрансферазным центром, который катализирует образование пептидной связи.
Долгое время считали, что трансляцию обеспечивают рибосомные белки, а рРНК служит лишь каркасом для их сборки. Однако в настоящее время доказано, что роль главного катализатора трансляции играет рРНК, а белки выполняют структурную функцию.
Между субъединицами рибосомы существует разделение труда. Малая субъединица содержит декодирующий центр, который обеспечивает взаимодействие между мРНК и тРНК. Большая субъединица содержит пептидилтрансферазный центр. В организации декодирующего центра участвуют 16S рРНК и рибосомные белки, в то время как пептидилтрансферазный центр образован только 23S рРНК. Последовательность Шайна-Дальгарно (J. Shine, L. Dalgarno), предшествующая стартовому кодону мРНК, спаривается с комплементарной последовательностью на З"-конце 16S рРНК. Антикодоновый конец тРНК также взаимодействует с 16S рРНК, тогда как акцепторный конец тРНК взаимодействует с 23S рРНК.
Рибосома шаг за шагом образует пептидные связи в направлении от N-конца к С-концу. Для инициации полипептидной цепи у бактерий используется особая аминокислота - формилметионин, которая доставляется в рибосому с помощью специфической тРНК. Происходит спонтанная пептидилтрансферазная реакция: нуклеофильная а-аминогруппа аминоацил-тРНК атакует электрофильную карбонильную группу (*) в сложной эфирной связи между пептидом (или формил-метиониновой затравкой) и другой тРНК. Молекулы 16S рРНК и 23S рРНК образуют три сайта -Р, А и Е, каждый из которых представлен субсайтами в обеих субъединицах рибосомы. P-сайт (от англ. peptide) связывает пептидил-тРНК, A-сайт (от англ. amino acid) связывает аминоацил-тРНК, а Е-сайт (от англ. exit) связывает деацилированную тРНК.
Рабочий цикл рибосомы состоит из четырех этапов, или состояний.
1. В исходном состоянии Р/Р-А/А пептидил-тРНК находится в P-сайте, аминоацил-тРНК в A-сайте, Е-сайт свободен. Специфическая аминоацил-тРНК связывается с A-сайтом при помощи фактора элонгации EF-Tu. Для этого используется энергия гидролиза ГТФ. Тройственный комплекс аминоацил-тРНК/(ЕГ-Ти) × ГТФ прочно связывается с рибосомой только если антикодон комплементарен кодону в декодирующем субсайте А.
2. В «претранслокационном» состоянии Р/Р-А/А происходит пептидилтрансфераз-ная реакция. При этом аминокислота, находящаяся в A-сайте, образует связь с пептидом (или формилметионином, если цепь инициируется), который находится в P-сайте. В обоих случаях дипептид или полипептидная цепь, удлиннившаяся на один аминокислотный остаток, переносятся в А-сайт. Чтобы следующая молекула аминоацил-тРНК могла попасть в A-сайт, пептидил-тРНК должна освободить его и перейти в P-сайт. Этот процесс называется «транслокацией». При транслокации взаимодействующие друг с другом тРНК и мРНК перемещаются внутри рибосомы на расстояние до 50А.
3. В «гибридном транслокационном» состоянии Е/Р-Р/А связанный с пептидом конец тРНК перемещается на большой субъединице из А-субсайта в Р-субсайт, а акцепторный ССА-конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. Этот этап транслокации напоминает «эффект домино» и зависит от рРНК. Его механизм еще неизвестен.
4. В «однородном транслокационном» состоянии Е/Е-Р/Р антикодоновый конец тРНК, связанной с пептидом, перемещается на малой субъединице из А-субсайта в ее же Р-субсайт, а антикодоновый конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. В результате этого мРНК сдвигается в малой субъединице на один кодон. Теперь A-сайт может принять следующую молекулу аминоацил-тРНК, а деацилированная молекула тРНК может покинуть рибосому. Хотя этот этап транслокации зависит от рРНК, он ускоряется фактором элонгации EF-G, который использует энергию гидролиза ГТФ.
Действие многих антибиотиков (канамицина, неомицина, олеандомицина, стрептомицина, тетрациклина, хлорамфеникола и др.) основано на их связывании с факторами элонгации и сайтами, которые образует рРНК.
Структура рибосом эукариот
Рибосомы состоят из двух различных субчастиц, каждая из которых построена из рибосомной РНК и многих белков. Рибосомы и их субчастицы обычно классифицируют не по массам, а в соответствии с коэффициентами седиментации. Так. коэффициент седиментации полной эукариотической рибосомы составляет около 80 единиц Сведберга (80S), а коэффициент седиментации ее субчастиц составляет 40S и 60S.
Меньшая 40S-субчастица состоит из одной молекулы 18S-рРНК и 30-40 белковых молекул. Большая 60S-субчастица содержит три типа рРНК с коэффициентами седиментации 5S, 5,8S и 28S и 40-50 белков (например, рибосомы гепатоцитов крысы включают 49 белков). В присутствии мРНК (mRNA) субчастицы объединяются с образованием полной рибосомы, масса которой примерно в 650 раз больше массы молекулы гемоглобина. Рибосомы имеют диаметр 20-200 нм и их можно видеть в электронный микроскоп. Структурная организация рибосом полностью не выяснена. Однако известно, что молекулы мРНК проходит через щель около характерной структуры в виде «рога» на малой субчастице, причем эта щель ориентирована как раз в промежуток между двумя субчастицами. тРНК также связываются вблизи этого участка. Для сравнения на схеме в том же масштабе показана молекула тРНК.
В клетках эукариот рибосомы формируются в ядрышке, где на ДНК синтезируется р-РНК, к которой затем присоединяются белки. Субчастицы рибосомы выходят из ядра в цитоплазму, и здесь завершается формирование полноценных рибосом. В цитоплазме рибосомы свободно находятся в цитоплазматическом матриксе (гиалоплазме) или прикрепляются к внешним мембранам ядра и эндоплазматической сети. Свободные рибосомы синтезируют белки для внутренних нужд клетки. Рибосомы на мембранах образуют комплексы – полирибосомы, которые синтезируют белки, поступающие через эндоплазматическую сеть в аппарат Гольджи и затем секретируемые клеткой. Количество рибосом в клетке зависит от интенсивности биосинтеза белка – их больше в клетках активно растущих тканей (меристем растений, зародышей и т. п.). В хлоропластах и митохондриях есть свои собственные мелкие рибосомы, они обеспечивают этим органоидам автономный (независимый от ядра) биосинтез белков.
Каждая рибосома состоит из двух субчастиц - большой и малой. Рибосомы состоят из примерно равных (по массе) количеств РНК и белка (т.е. представляют собой рибонуклеопротеиновые частицы). Входящая в их состав РНК, называемая рибосомной РНК (рРНК), синтезируется в ядрышке. Вместе те и другие образуют сложную трехмерную структуру, обладающую способностью к самосборке. Во время синтеза белка на рибосомах аминокислоты, из которых строится полипептидная цепь, последовательно одна за другой присоединяются к растущей цепи. Рибосома служит местом связывания для молекул, участвующих в синтезе, т. е. таким местом, где эти молекулы могут занять по отношению друг к другу совершенно определенное положение.
В синтезе участвуют: матричная РНК (мРНК), несущая генетические инструкции от ядра клетки, транспортная РНК (тРНК), доставляющая к рибосоме требуемые аминокислоты, растущая полипептидная цепь, а также ряд факторов, ответственные за инициацию, элонгацию и терминацию цепи. В эукариотических клетках отчетливо видны две популяции рибосом - свободные рибосомы и рибосомы, присоединенные к эндоплазматическому ретикулуму. Строение тех и других идентично, но часть рибосом связана с эндоплазматическим ретикулоумом через белки, которые они синтезируют. Такие белки обычно секретируются. Примером белка, синтезируемого свободными рибосомами, может служить гемоглобин, образующийся в молодых эритроцитах. В процессе синтеза белка рибосома перемещается вдоль нитевидной молекулы мРНК. Процесс идет более эффективно, когда вдоль мРНК перемещается не одна рибосома, а одновременно много рибосом, напоминающих в этом случае бусины на нитке. Такие цепи рибосом называются полирибосомами или полисомами. На эндоплазматическом ретикулуме полисомы обнаруживаются в виде характерных завитков.
Рибосомный синтез белка-многоэтапный процесс. Первая стадия (инициация) начинается с присоединения матричной РНК (мРНК) к малой рибосомной субчастице, не связанной с большой субчастицей. Характерно, что для начала процесса необходима именно диссоциированная рибосома. К образовавшемуся т. наз. инициаторному комплексу присоединяется большая рибосомная субчастица. В стадии инициации участвуют спец. инициирующий кодон (см. Генетический код), инициаторная транспортная РНК (тРНК) и специфич. белки (т. наз. факторы инициации). Пройдя стадию инициации, рибосома переходит к последоват. считыванию кодонов мРНК по направлению от 5"- к 3"-концу, что сопровождается синтезом полипептидной цепи белка, кодируемого этой мРНК В этом процессе рибосома функционирует как циклически работающая мол. машина.
Рабочий цикл рибосомы при элонгации состоит из трех тактов: 1) кодонзависимого связывания аминоацил-тРНК (поставляет аминокислоты в рибосому), 2) транспептидации-переноса С-конца растущего пептида на аминоацил-тРНК, т.е. удлинения строящейся белковой цепи на одно звено, 3) транслокации-перемещения матрицы (мРНК) и пептидил-тРНК относительно рибосомы и переход рибосомы в исходное состояние, когда она может воспринять след. аминоацил-тРНК. Когда рибосома достигнет специального терминирующего кодона мРНК, синтез полипептида прекращается. При участии специфич. белков (т. наз. факторов терминации) синтезир. полипептид освобождается из рибосомы. После терминации рибосома может повторить весь цикл с др. цепью мРНК или др. кодирующей последовательностью той же цепи.
В клетках с интенсивной секрецией белка и развитым эндоплазматич. ретикулумом значит. часть цитоплазматической рибосомы прикреплена к его мембране на поверхности, обращенной к цитоплазме. Эти рибосомы синтезируют полипептиды, которые непосредственно транспортируются через мембрану для дальнейшей секреции. Синтез полипептидов для внутриклеточных нужд происходит в основном на свободных (не связанных с мембраной) рибосомах цитоплазмы. При этом транслирующие рибосомы не равномерно диспергированы в цитоплазме, а собраны в группы. Такие агрегаты рибосом представляют собой структуры, где мРНК ассоциирована со многими рибосомами, находящимися в процессе трансляции; эти структуры получили назв. полирибосом или полисом.
При интенсивном синтезе белка расстояние между рибосомами вдоль цепи мРНК в полирибосоме может быть предельно коротким, т.е. рибосомы находятся почти вплотную друг к другу. Рибосомы, входящие в полирибосомы, работают независимо и каждая из них синтезирует полную полипептидную цепь.
Отличия в строении рибосом прокариотов и эукариотов
Прокариотическая клетка содержит несколько тысяч рибосом, в эукариотической клетке их в десятки раз больше. Рибосомы про- и эукариот отличаются по размерам (у прокариот они мельче, чем у эукариот), но принцип их строения одинаков. Состоят рибосомы из двух частей: большой и малой субъединиц. В их состав кроме белков входят РНК. Эти РНК получили название рибосомных, рРНК.
Величину рибосом и составляющих их частей принято указывать в специальных единицах - S (Сведберг). S - это коэффициент седиментации, который характеризует скорость перемещения молекул или частиц в центробежном поле при центрифугировании. Скорость перемещения зависит от массы частиц, их размеров и формы. Величина рибосом прокариот и эукариот - 70S и 80S соответственно.
В рибосомы прокариот входит три разных вида молекул рРНК (16S рРНК - в малую; 23S рРНК и 5S рРНК - в большую субъединицы) и 55 различных белков (21 - в малую и 34 - в большую субъединицы). В состав эукариотических рибосом входят четыре вида молекул рРНК (18S рРНК - в малую; 28S рРНК, 5.8S рРНК и 5S рРНК - в большую субъединицы) и около 80 белков. В митохондриях и хлоропластах также обнаружены рибосомы. Они характеризуются теми же свойствами и параметрами, что и рибосомы прокариот.
Молекулы рРНК взаимодействуют друг с другом и с белками, образуя компактные структуры - субъединицы рибосом. У эукариот соединение рРНК с рибосомными белками происходит в ядрышке. В центре ядрышка расположен участок хромосомы, в котором находятся гены рибосомных РНК. Синтезированные рРНК соединяются с рибосомными белками, которые поступили через ядерные поры из цитоплазмы, где они были синтезированы на уже существовавших рибосомах. Они соединяются с молекулами рРНК, образуя субъединицы рибосом. Готовые субъединицы через поры выходят в цитоплазму, где будут участвовать в синтезе белка.
Таким образом, ядрышко - это не только место синтеза рибосомных РНК, но и место сборки субъединиц рибосом. Рибосомы нужны в огромных количествах, поскольку в клетке постоянно идут процессы синтеза белка. Поэтому на хромосомах в тех местах, где расположены гены рРНК, находится громадное скопление молекул: синтезируемые рРНК, пришедшие из цитоплазмы рибосомные белки, собираемые и готовые суъединицы рибосом. Понятно, почему ядрышко является самой плотной частью ядра и клетки. Размеры ядрышка зависят от функционального состояния клеток. Если в клетке активно идут процессы биосинтеза белков, ядрышко может занимать до 25% от объема ядра.
Ядрышко образуется на тех хромосомах, где есть гены рРНК. Эти участки хромосом называются ядрышковыми организаторами. Например, у человека десять хромосом способны образовывать ядрышки. Каждый ядрышковый организатор представляет собой огромную хроматиновую петлю, так как содержит несколько десятков и даже сотен одинаковых последовательностей - генов рРНК. Эти последовательности расположены друг за другом и синтез рРНК идет одновременно со всех копий. Таким образом увеличивается интенсивность синтеза рРНК, на долю которой приходится более 90 % всей РНК клетки. Ядрышки, образованные разными хромосомами, очень часто сливаются друг с другом. В ядрах клеток человека обычно наблюдают одно, два или три ядрышка.
При начале трансляции малая субъединица рибосомы связывается с определенным участком иРНК, к ним присоединяется тРНК с аминокислотой, а затем с этим комплексом связывается большая субъединица. После этого рибосома готова к выполнению своей функции - синтезу белка. Белки рибосом способны выполнять свои функции только в составе рибосомы -только в комплексе с рРНК и другими рибосомными белками они приобретают небходимую конформацию.
Эукариотная транскрипция разделена с трансляцией в пространстве и времени. Транскрипция вместе с процессингом РНК происходят в нуклеоплазме, а трансляция, в зависимости от типа клеток, осуществляется преимущественно в цитозоле или на шероховатом эндогшазматическом ретикулуме (англ. rough endoplasmic reticulun, RER). Интегральные белки встраиваются в мембрану RER котрансляционно, а секретируемые белки выделяются в полость цистерны RER через тороидальный переходник между выходным порталом рибосомы и мембранным транслоконом (его образует белок Sec61).
У прокариотов не существует пространственно-временной изоляции процессов транскрипции и трансляции. Цитоплазматические рибосомы присоединяются к 5"-концу мРНК еще до завершения образования короткоживущего транскрипта. Котрансляционная инсерция интегральных белков известна только на примере «шероховатых тилакоидов» цианобактерий. Гидрофобные белки при помощи SRP-частиц презентируются транс локону - компоненту генеральной системы секреции Sec.
Транспортная РНК, напоминает в развернутой форме клеверный лист. Аминокислота прикреплена к “черешку клеверного листа”, а на вершине листа находится триплет, взаимодействующий с кодоном в иРНК - антикодон. Роль "заглавной буквы" при трансляции аминокислотной последовательности у прокариот выполняет измененная форма аминокислоты метионина - формилметионин. Ей соответствует кодон АУГ. После завершения синтеза полипептидной цепи формилметионин отщепляется и в готовом белке отсутствует. В том случае, когда триплет АУГ стоит внутри гена, он кодирует неизмененную аминокислоту метионин.
Если кодон и антикодон комплементарны друг другу, то рибосома передвигается относительно иРНК, и следующий кодон становится доступным для взаимодействия со следующей тРНК. Происходит отсоединение первой аминокислоты от первой тРНК и присоединение ее к аминокислоте, которую принесла вторая тРНК. Во время передвижения рибосомы относительно иРНК первая тРНК, свободная от аминокислоты, покидает рибосому. Вторая тРНК остается, соединенная с пептидом из двух аминокислотных остатков, и в рибосому входит третий кодон иРНК для взаимодействия с очередной тРНК и т.д.
Когда в рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА), ни одна тРНК не может занять место напротив него, так как не существует тРНК с антикодонами, комплементарными этим последовательностям. Полипептидной цепи не к чему присоединиться и она покидает рибосому. Синтез белка завершен. Таким образом, рибосома соединяет в одном месте участников трансляции: иРНК и аминокислоты в комплексе с тРНК, при этом молекулы РНК так ориентированы относительно друг друга, что становится возможным кодон-антикодоновое взаимодействие. Образование пептидной связи контролируется правильностью кодон-антикодонового взаимодействия. Рибосома осуществляет образование пептидной связи и перемещение относительно иРНК.
Молекула информационной РНК взаимодействует не с одной рибосомой, а с несколькими. Каждая рибосома проходит весь путь от "заглавного" кодона до терминирующего, синтезируя одну молекулу белка. Чем больше рибосом пройдет по иРНК, тем больше молекул белка будет синтезировано. Молекула информационной РНК с несколькими рибосомами похожа на нитку бус и называется полирибосомой, или полисомой.
Рибосомы являются важнейшими органоидами клетки, так как на них протекает процесс трансляции - синтез полипептида на матричной РНК (мРНК). Другими словами, рибосомы служат местом белкового синтеза .
Строение рибосом
Рибосомы относятся к немембранным органоидам. Они очень мелкие (около 20 нм), но многочисленные (тысячи и даже миллионы на клетку), состоят из двух частей – суб ъединиц . В состав субчастиц входят рибосомальные РНК (рРНК) и рибосомные белки, т. е. рибосомы по химическому составу являются рибонуклеопротеи д ами . Однако в них также присутствует небольшое количество низкомолекулярных соединений. Из-за многочисленности рибосом, рРНК составляет более половины от всей РНК клетки.
Одну из субъединиц называют «малой», вторую – «большой».
В собранной из субъединиц рибосоме выделят два (по одним источникам) или три (по другим) участка, которые называют сайтами . Один из участков обозначают A (aminoacyl) и называют аминоацильным, второй - P (peptidyl) - пептидильный. Данные сайты являются основными каталитическими центрами протекающих на рибосомах реакций. Третий участок обозначают E (exit), через него освободившаяся от синтезируемого полипептида транспортная РНК (тРНК), покидает рибосому.
Кроме перечисленных сайтов на рибосомах есть другие участки, используемые для связывания различных ферментов.
Когда субъединицы диссоциированы (разъединены) специфичность сайтов теряется, т. е. они определяются сочетанием соответствующих областей обеих субъединиц.
Отличие рибосом прокариот и эукариот
Соотношение по массе белков и РНК в рибосоме примерно поровну. Однако у прокариот белков меньше (около 40%).
Размеры как самих рибосом, так и субъединиц выражают в скорости их седиментации (осаждения) при центрифугировании. При этом S обозначает константу Сведберга - единицу, характеризующую скорость оседания в центрифуге (чем больше S, тем быстрее частица осаждается, а значит тяжелее). У прокариот рибосомы имеют размер в 70S, а у эукариот - в 80S (т. е. они тяжелее и крупнее). При этом субъединицы прокариотических рибосом имеют значения 30S и 50S, а эукариотических - 40S и 60S. Размеры рибосом в митохондриях и хлоропластах эукариот сходны с прокариотическими (хотя имеют определенную вариабельность по размерам), что может указывать на их происхождение от древних прокариотических организмов.
У прокариот в состав большой субъединицы рибосом входит две молекулы рРНК и более 30 молекул белка, в состав малой - одна молекула рРНК и около 20 белков. У эукариот в субъединицах больше молекул белка, а также в большой субъединице три молекулы рРНК. Составляющие рибосому белки и молекулы рРНК обладают способностью к самосборке и в итоге образуют сложную трехмерную структуру. Структуру рРНК поддерживают ионы магния.
Синтез рРНК
У эукариот в состав рибосом входят 4 вида рРНК. При этом три образуются из одного транскрипта-предшественника - 45S рРНК. Он синтезируется в ядрышке (на петлях хромосом его формирующем) при помощи РНК-полимеразы-1. Гены рРНК имеют много копий (десятки и сотни) и обычно располагаются на концах разных пар хромосом. После синтеза 45S рРНК разрезается на 18S, 5.8S и 28S рРНК, каждая из которых подвергается тем или иным модификациям.
Четвертый вид рРНК синтезируется вне ядрышка с помощью фермента РНК-полимеразы-3. Это 5S РНК, которая после синтеза не нуждается в .
Третичная структура рРНК в составе рибосом очень сложная и компактная. Она служит каркасом для размещения рибосомных белков, которые выполняют вспомогательные функции для поддержания структуры и функциональности.
Функция рибосом
Функционально рибосомы являются местом связывания молекул, участвующих в синтезе (мРНК, тРНК, различные факторы). Именно в рибосоме молекулы могут занять друг по отношению к другу такое положение, которое позволит быстро протечь химической реакции реакции.
В эукариотических клетках рибосомы могут находиться свободно в цитоплазме или быть прикрепленными с помощью специальных белков к ЭПС (эндоплазматическая сеть, она же ЭР - эндоплазматический ретикулум).
В процессе трансляции рибосома перемещается по мРНК. Часто по одной нитевидной мРНК двигаются несколько (или множество) рибосом, образуя так называемую полисому (полирибосому).
| Наименование параметра | Значение |
| Тема статьи: | Рибосомы |
| Рубрика (тематическая категория) | Образование |
Строение рибосомы: 1 - большая субъединица; 2 - малая субъединица.
Рибосомы - немембранные органоиды, диаметр примерно 20 нм. Рибосомы состоят из двух субъединиц - большой и малой, на которые могут диссоциировать. Химический состав рибосом - белки и рРНК. Молекулы рРНК составляют 50–63% массы рибосомы и образуют ее структурный каркас. Различают два типа рибосом: 1) эукариотические (с константами седиментации целой рибосомы - 80S, малой субъединицы - 40S, большой - 60S) и 2) прокариотические (соответственно 70S, 30S, 50S).
В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа - 3 молекулы рРНК и около 55 молекул белка. Во время биосинтеза белка рибосомы могут ʼʼработатьʼʼ поодиночке или объединяться в комплексы - полирибосомы (полисомы) . В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).
Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Рибосомы - понятие и виды. Классификация и особенности категории "Рибосомы" 2017, 2018.
Существуют два пути узнавания инициирующего кодона Терминальная инициация (характерна для эукариот), представляет процесс, при котором рибосома точно присоединяется к модифицированному (кэп) 5’-концу иРНК и перемещается по ней до тех пор, пока не обнаружит... .
Рибосомы состоят из двух субъединиц неравного размера - большой и малой, на которые они могут диссоциировать. В состав рибосом входят белки и рибосомальные РНК (рРНК). В зависимости от локализации в клетке, различают © свободные рибосомы - рибосомы, находящиеся в... .
Рибосома - это округлая рибонуклеопротеиновая частица диаметром 20-30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки... .
Вакуоли и сферосомы растительных клеток Секреторные везикулы и гранулы Этот тип одномембранных органоидов связан с экзоцитозом – синтезом и выделением веществ из клетки. Различают две разновидности экзоцитоза: секрецию и экскрецию.... .
Ядрышко Ядрышко представляет собой хорошо заметную округлую структуру, находится внутри ядра. Это место образования рибосом. В ядре может быть одно или несколько ядрышек. Оно интенсивно окрашивается, потому что содержит большое количество ДНК и РНК. В ядрышке имеется... .
Рибосомы – немембранные органеллы клетки, функция которых заключается в биосинтезе белка. Каждая рибосома состоит из двух субъединиц – большой и малой, построенных из рибосомной РНК и белков. По массе рибосомной РНК в рибосоме больше – из нее построены каркасы большой... .
По количеству преобладают палочки (120-130 млн). Слой фоторецепторных нейронов - самый широкий, содержит фоторецепторные клетки - это первично-чувствующие клетки, у них есть аксон, который соединяется с дендритами ассоциативных нейронов и периферический отросток... .
Каждая клетка любого организма имеет сложную структуру, включающую в себя множество компонентов.
Вкратце о строении клетки
Она состоит из мембраны, цитоплазмы, органоидов, которые в них расположены, а также ядра (кроме прокариотов), в котором находятся молекулы ДНК. Кроме того, над мембраной имеется дополнительная защитная структура. В животных клетках во всех остальных - У растений она состоит из целлюлозы, у грибов - из хитина, у бактерий - из муреина. Мембрана состоит из трех слоев: двух фосфолипидных и белкового между ними.
В ней есть поры, благодаря которым осуществляется перенос веществ внутрь и наружу. Возле каждой поры расположены специальные транспортные белки, которые пропускают в клетку только определенные вещества. Органоидами животной клетки являются:
Рибосома - это что?
Раз уж мы говорим о ней в данной статье, то вполне логично задать такой вопрос. Рибосома - это органоид, который может быть расположен на внешней стороне стенок комплекса Гольджи. Нужно уточнить еще, что рибосома - это органоид, который содержится в клетке в очень больших количествах. В одной может находиться до десяти тысяч.

Где находятся данные органоиды?
Итак, как уже говорилось, рибосома - это структура, которая находится на стенках комплекса Гольджи. Также она может свободно передвигаться по цитоплазме. Третий вариант, где может располагаться рибосома - мембрана клетки. И те органоиды, которые находятся в этом месте, практически не покидают его и являются стационарными.
Рибосома - строение
Как же выглядит данная органелла? Она похожа на телефон с трубкой. Рибосома эукариот и прокариот состоит из двух частей, одна из которых больше, другая - меньше. Но эти две ее составляющие не соединяются вместе, когда она находится в спокойном состоянии. Это происходит только тогда, когда рибосома клетки непосредственно начинает выполнять свои функции. О функциях мы поговорим позже. Рибосома, строение которой описывается в статье, также имеет в своем составе информационную РНК и Данные вещества необходимы для того, чтобы записывать на них информацию о нужных клетке белках. Рибосома, строение которой мы рассматриваем, не имеет собственной мембраны. Ее субъединицы (так называются две ее половины) ничем не защищены.

Какие функции выполняет в клетке данный органоид?
То, за что отвечает рибосома, - синтез белка. Он происходит на основе информации, которая записана на так называемой матричной РНК (рибонуклеиновой кислоте). Рибосома, строение которой мы рассмотрели выше, объединяет свои две субъединицы только на время синтеза белка - процесса под названием трансляция. Во время данной процедуры синтезируемая полипептидная цепь находится между двумя субъединицами рибосомы.
Где они формируются?
Рибосома - органоид, который создается ядрышком. Данная процедура происходит в десять этапов, на протяжении которых постепенно формируются белки малой и большой субъединиц.
Каким образом происходит формирование белков?
Биосинтез белков происходит в несколько этапов. Первый из них - это активация аминокислот. Всего их существует двадцать, при комбинировании их разными методами можно получить миллиарды различных белков. На протяжении данного этапа из аминокислот формируется аминоалиц-т-РНК. Данная процедура невозможна без участия АТФ (аденозинтрифосфорной кислоты). Также для осуществления этого процесса необходимы катионы магния.

Второй этап - полипептидной цепи, или процесс объединения двух субъединиц рибосомы и поставка к ней необходимых аминокислот. В данном процессе также принимают участие ионы магния и ГТФ (гуанозинтрифосфат). Третий этап называется элонгацией. Это непосредственно синтез полипептидной цепи. Происходит методом трансляции. Терминация - следующий этап - это процесс распада рибосомы на отдельные субъединицы и постепенное прекращение синтеза полипептидной цепочки. Далее идет последний этап - пятый - На этой стадии из простой цепи аминокислот формируются сложные структуры, которые уже и представляют собой готовые белки. В данном процессе участвуют специфические ферменты, а также кофакторы.
Структура белка
Так как рибосома, строение и функции которой мы разобрали в этой статье, отвечает за синтез белков, то давайте рассмотрим подробнее их структуру. Она бывает первичной, вторичной, третичной и четвертичной. - это определенная последовательность, в которой располагаются аминокислоты, формирующие данное органическое соединение. представляет собой сформированные из полипептидных цепочек альфа-спирали и бета-складки. Третичная структура белка предусматривает определенную комбинацию альфа-спиралей и бета-складок. Четвертичная же структура заключается в формировании единого макромолекулярного образования. То есть комбинации альфа-спиралей и бета-структур формируют глобулы либо фибриллы. По этому принципу можно выделить два типа белков - фибриллярные и глобулярные.

К первым относятся такие, как актин и миозин, из которых сформированы мышцы. Примерами вторых могут служить гемоглобин, иммуноглобулин и другие. напоминают собой нить, волокно. Глобулярные больше похожи на клубок сплетенных между собой альфа-спиралей и бета-складок.
Что такое денатурация?
Каждый наверняка слышал это слово. Денатурация - это процесс разрушения структуры белка - сначала четвертичной, затем третичной, а после - и вторичной. В некоторых случаях происходит и ликвидация первичной структуры белка. Данный процесс может происходить вследствие воздействия на данное органическое вещество высокой температуры. Так, денатурацию белка можно наблюдать при варке куриных яиц. В большинстве случаев этот процесс необратим. Так, при температуре выше сорока двух градусов начинается денатурация гемоглобина, поэтому сильная гипертермия опасна для жизни. Денатурацию белков до отдельных нуклеиновых кислот можно наблюдать в процессе пищеварения, когда с помощью ферментов организм расщепляет сложные органические соединения на более простые.

Вывод
Роль рибосом очень сложно переоценить. Именно они являются основой существования клетки. Благодаря данным органоидам она может создавать белки, которые ей необходимы для самых разнообразных функций. формирующиеся рибосомами, могут играть защитную роль, транспортную, роль катализатора, строительного материала для клетки, ферментативную, регуляторную (многие гормоны имеют белковую структуру). Поэтому можно сделать вывод, что рибосомы выполняют одну из самых важных функций в клетке. Поэтому их и так много - клетке всегда нужны продукты, синтезируемые данными органоидами.
Рибосома - это маленькая электронно-плотная частица, образованная связанными между собой молекулами рРНК и белками, которые формируют сложное надмолекулярное соединение - рибонуклеопротеидный комплекс.
В рибосомах белки и молекулы рРНК находятся примерно в равных весовых отношениях. В состав цитоплазматических рибосом эукариот входят четыре молекулы рРНК, различающиеся по молекулярной массе. Количество органелл в клетке весьма разнообразно: тысячи и десятки тысяч. Рибосомы могут быть связаны с ЭПС или находиться в свободном состоянии.
Рибосома представляет собой сложное органическое соединение, формирующее компактную органеллу, способную считывать информацию с цепей иРНК и, используя ее, синтезировать полипептидные цепочки.
Рибосома расшифровывает информационный код, содержащийся в иРНК, который составлен четырьмя видами нуклеотидов. Три нуклеотида, располагаясь в различных последовательностях, несут информацию о двадцати аминокислотах. Рибосома, по сути дела, исполняет роль переводчика этой информации. Эта задача разрешается с помощью тРНК и ферментов, синтезирующих полипептидные цепочки. Такие ферменты называются аминоацил-тРНК-синтетазами. Число аминоацил-тРНК-синтетаз определяется разнообразием аминокислот, так как каждой аминокислоте соответствует свой фермент. Таким образом, в каждой рибосоме не менее 20 видов таких ферментов.
Рибосома состоит из большой и малой субъединиц. Каждая из субъединиц построена из рибонуклеопротеидного тяжа, где рРНК взаимодействует со специальными белками и образует тело рибосомы. Рибосомы образуются в ядрышке или матриксе митохондрий. Синтез полипептидных цепочек, осуществляемый рибосомами, называется трансляцией рРНК - это основа для формирования рибосом. Малая субъединица рибосомы образована одной молекулой рРНК и примерно 30 белками. В большую субъединицу встроена одна длинная рРНК и две коротких. С ними связаны 45 молекул белков.
тРНК - это небольшие молекулы, состоящие из 70…90 нуклеотидов, которые имеют форму листа клевера. тРНК доставляет аминокислоты к рибосомам. Каждая молекула тРНК имеет акцепторный конец, к которому присоединяется активированная аминокислота. Аминокислоты прикрепляются к последовательности трех нуклеотидов, комплементарных (соответствующих) нуклеотидам кодона в иPHК - антикодону.
Различают цитоплазматические (свободные и связанные) и митохондриальные рибосомы. Цитоплазматические и митохондриальные рибосомы значительно отличаются друг от друга по химическому составу, размерам и происхождению.
При электронной микроскопии обнаруживают как единичные рибосомы, так и их комплексы (полисомы). Вне синтеза субъединицы рибосом располагаются отдельно друг от друга. Субъединицы объединяются в момент трансляции информации с иРНК. При этом трансляция информации с одной молекулы иРНК осуществляют несколько рибосом (от 5…6 до нескольких десятков). Такие рибосомы чаще всего формируют так называемые полисомы - рыхлый конгломерат рибосом, располагающийся цепочкой по ходу иРНК. Это позволяет синтезировать с одной молекулы иРНК сразу несколько полипептидных цепочек.
Вне трансляции субъединицы рибосом могут распадаться и вновь объединяться. Этот процесс находится в динамическом равновесии. Процесс трансляции запускается со сборки активной рибосомы и обозначается как инициация трансляции. В собранной рибосоме имеются активные центры. Такие центры располагаются на контактирующих поверхностях обеих субъединиц. Между малой и большой субъединицами располагается серия углублений. В этих полостях находятся: иРНК, тРНК и синтезируемый пептид (пептидил-тРНК). Зоны, связанные с синтетическими процессами, формируют следующие активные центры:
- центр связывания иРНК (М-центр);
- пептидильный центр (П-центр), на котором происходит инициация и окончание считывания информации, а в процессе синтеза полипептида на нем находится полипептидная цепочка;
- аминокислотный центр (A-центр), место связывания с очередной тРНК;
- пептидилтрансферазный центр (ПТФ-центр). Здесь происходит катализ синтеза полипептида и синтезируемая молекула удлиняется на еще одну аминокислоту.
На малой субъединице расположен М-центр, основная часть A-центра и небольшой участок П-центра. На большой субъединице можно найти остальные части А- и П-центров, а также ПТФ-центр.
Трансляция начинается со стартового кодона - триплета аденин-урацил-гуанин, расположенного в 5′-конце иРНК. Он присоединяется к малой субъединице на уровне П-центра будущей рибосомы. Затем происходит объединение комплекса с большой субъединицей. Этот процесс активируют или, наоборот, блокируют белковые факторы. С момента формирования рибосома прерывисто, триплет за триплетом движется вдоль молекулы и РНК, что сопровождается ростом полипептидной цепочки. Число аминокислот в таком белке равно числу триплетов иРНК.
Процесс трансляции предполагает цикл близких событий и называется элонгацией - удлинение пептидной цепочки. Сигналом для прекращения трансляции служит появление в иРНК одного из «бессмысленных» кодонов (УАА, УАГ, УГА). Эти кодоны узнает один из двух факторов терминации. Они активируют гидролазную активность пептидилтрансферазного центра, что сопровождается отщеплением образованного полипептида, распадом рибосомы на субъединицы и прекращением синтеза.
Свободные рибосомы распределены в матриксе цитоплазмы. Они находятся либо в виде субъединиц и не участвуют в трансляции, либо «считывают» информацию, образуя полипептидные цепочки белков матрикса цитоплазмы и ядра, цитоскелета клетки и т. д.
Связанные рибосомы - это такие рибосомы, которые прикреплены к мембранам гр. ЭПС или к наружной мембране ядерной оболочки. Происходит это только в момент синтеза полипептидных цепочек белков, формирующих секреторные гранулы цитолеммы, лизосом, ЭПС, комплекса Гольджи и др.
Синтез белковых молекул происходит непрерывно и идет с большой скоростью: в одну минуту образуются от 50 до 60 тыс. пептидных связей. За одну секунду рибосома эукариот считывает информацию с 2…15 кодонов (триплетов) иРНК. Синтез одной молекулы крупного белка (глобулина) длится около 2 мин. У бактерий этот процесс идет гораздо быстрее.
Таким образом, рибосомы - это органеллы, обеспечивающие анаболические процессы в клетке, а именно синтез полипептидных цепочек белков.
В слабо специализированных и быстро растущих клетках в основном обнаруживают свободные рибосомы. В специализированных клетках рибосомы располагаются в составе гр. ЭПС. Содержание РНК и соответственно степень белковых синтезов соотносится с количеством рибосом. Это сопровождается склонностью к базофилии цитоплазмы, то есть способностью окрашиваться основными красителями.
В клетках некоторых типов цитоплазма более базофильна, чем в других. Базофилия может быть диффузной или локальной. С помощью электронной микроскопии установлено, что локальная базофилия создается гр. ЭПС, а именно прикрепленными к ее мембранам рибосомами. Примерами подобной, фокусной базофилии служат: цитоплазма нейрона, базальный полюс железистого эпителия концевых отделов экзокринной части поджелудочной железы, белковопродутдирующие клетки слюнных желез. Диффузная базофилия обусловлена свободными рибосомами. Базофилию выявляют и в случае накопления в цитоплазме включений или большого количества лизосом, имеющих кислое содержимое. В этих случаях видна базофильно окрашенная зернистость.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .